Chapter 5: Hypernotes
![]() WWW☜ 5:1
WWW☜ 5:1
Act II, Scene II of Siegfried, in the English translation by Andrew Porter, performed by the English National Opera. The libretto here is quoted from the complete recording in August 1973, conducted by Sir Reginald Goodall at the London Coliseum (EMI SLS875, OC 191 02473-02477).
Wagner himself does not specify that the heart is on the left, and he may only, for instance, have meant that the heart was in the chest. The original words, with a very literal translation, are:
“Doch heiße mich das:
hat der Wurm ein Herz?
“Ein grimmiges, hartes Herz!
“Das sitzt ihm doch
wo es jedem schlägt,
trag’ es Mann oder Thier?
“Gewiß, Knabe,
da führt’s auch der Wurm;” (Wagner, 1888):
“Tell me this:
has the dragon a heart?"
"A grim, hard heart!"
“That must be in the same place
as it beats for everyone,
be they man or beast?"
“Certainly, ...,
that’s where the dragon has it as well...”
A detail to remember in speculating about the heart of Fafner the dragon is that earlier in the ring cycle Fafner is one of the two giants who steal the gold, and that Fafner uses the magical Tarnhelm to transform himself into Fafner the dragon to guard the stolen hoard. It is probable therefore that his internal organs will correspond quite closely to the internal organs of the giant Fafner, and therefore the heart will be on the left.
![]() WWW☜ 5:2
WWW☜ 5:2
The typical Western forms of dragon are found, for instance, in Carpaccio’s paintings of the life of St George in the Scuola di San Giorgio degli Schiavoni in Venice, and in Raphael's drawing in the Uffizi (Altmann, 1992 p.42), but also in innumerable books for children, and one can be seen, for instance, on the cover of J.K. Rowling’s Harry Potter and the Goblet of Fire. Portrayals of dragons go back to the earliest recorded civilisations and seem to be world-wide in their distribution. The determining feature is the inclusion of body parts from several or many different animals, as in an early Sumerian dragon with the claws and wings of an eagle and the hind legs of a lion (Elliot Smith, 1919 p.79, figure 1). An ancient Chinese description of a dragon describes its “nine resemblances” – “His horns resemble those of a stag, his head that of a camel, his eyes those of a demon, his neck that of a snake his belly that of a clam, his scales those of a carp, his claws those of an eagle, his soles those of a tiger, his ears those of a cow” (Elliot Smith, 1919 p.81).
![]() WWW☜ 5:3
WWW☜ 5:3
In Beowulf, the eponymous hero, Beowulf, fights the dragon Grendel bare-handed and mortally wounds him by ripping off his arm and shoulder, which are carried back to the great hall, Heorot, and put on display. While Grendel fled to his den to die, “his right hand [swīðre hand] stayed behind him in Heorot,/ evidence of his miserable overthrow/ as he dived into murk on the mere-bottom” (Heaney, 1999 p.67).

I could not resist including the illustration of the Burroughs-Wellcome advert in this book, the writing of which was supported by a prize from the Wellcome Trust, whose wealth mainly has come from sales of that and a wealth of other drugs. It therefore seems an appropriate tribute to include the illustration. However when I originally found the picture of the painting, I was somewhat suspicious of it, as can be seen by my comment about the Valentine's heart. Subsequently I found the wonderful monograph by Vinken (1999a) entitled The shape of the heart which asks precisely where, when, and why modern icons of the heart, such as ♡❤ ♣, are 'heart-shaped' when the real human heart looks nothing like it. The brief answers would seem to be, 'in Northern Italy', 'at the beginning of the fourteenth century', and 'as the result of an error in understanding Aristotle'. The palaeolithic cave painting in the advertisement is actually at Pindal in Spain, where it was discovered by the French abbe Henri Breuil. In his original account he thought that the mark was the animal's ear, and in his 1911 report he describes how “a large, more or less heart-shaped mark ... represents the ear flap”. Apparently a group of elephant hunters then suggested to him that the mark was the heart, principally because of a painting of another elephant, in Southern France, where the beast is being shot with three arrows above the left shoulder, where the heart would be. Subsequent workers have been sceptical that the mark is heart- shaped, or is indeed any particular shape, the whole thing being too vague to have a clear contour. As yet I have been unable to track down the elephant with the three arrows in the left shoulder, but this might just possibly be very slight evidence that indeed prehistoric man knew the heart was on the left side.
![]() WWW☜ 5:4
WWW☜ 5:4
The scarlet tree was the title of the second volume of Sitwell’s autobiography (Sitwell, 1946), from which the quotation is taken. The dust jacket of the first edition shows a beautiful representation of the vascular tree. Ziegler (1999 p.309) says that Sitwell commented that the title refers to the 'singing blood in the young veins' – although the figure on the dust jacket undoubtedly shows the arteries...
.![]() WWW☜ 5:5
WWW☜ 5:5
Although I have talked about the human body, most of these asymmetries can be found in other vertebrates as well, albeit sometimes in modified form. I remember once, in Andhra Pradesh in India, dissecting a cobra that had been caught in the hospital compound and being amazed at how even in this seemingly so symmetric an organism, the liver was as asymmetric as in humans, albeit being very much longer and thinner.
A potential confusion in thinking about the asymmetry of the body is that anatomists also talk about the right and left sides of the heart. The right ventricle is smaller and pumps blood at relatively low pressure to the lungs, whereas the left ventricle is much larger and pumps blood at higher pressure, through the aorta, to the rest of the body. Although the right ventricle happens to be closer to the right side of the body, it has not developed from the right side. Both ventricles actually develop in the embryo from a single tube, one from the front end of the tube and the other from the rear end. and it is of no real consequence that the larger of the two happens to be called the left ventricle. It might have made more sense and been less confusing if one were called the first and the other the second ventricle, reflecting the order in which blood passes through them. See Larsen (1998 p.105).
The asymmetry of the testicles is not only of size and height. The venous drainage of the right testicle goes directly into the inferior vena cava, whereas that of the left testicle enters into the left renal vein. The difference in size of the testicles is also reflected in the ovaries of women, and in both males and females the asymmetry is present at an extremely early stage of embryonic development, so early that the fetus can only be sexed by looking at the chromosomes (Mittwoch & Kirk, 1975) but nevertheless there is an asymmetry of the rudiments of the gonads.
Varicocoele in one large series was on the left side in all of 136 cases (Oster, 1971). Because of the different venous drainage of the right and left testicle, there can be reflux of blood back down the left testicular vein which can cause the varicocoele. The infertility is probably due to the increased blood flow raising the temperature of the scrotum and preventing spermatogenesis. Removal of the varicocoele often restores fertility. Very occasionally varicocoele occurs on the right hand side and it is then said always to be associated with situs inversus (Server et al., 1998).
![]() WWW☜ 5:7
WWW☜ 5:7
Although I comment mention "organisms such as sea-squirts, which are more primitive than vertebrates", it must be emphasised that this is merely short hand, since modern sea-squirts are neither our ancestors nor, in a strict sense, are more primitive than us, having been in existence for precisely the same amount of time (Gee, 2000). The loose terminology is however useful for a simple description.
Although having an internal skeleton solves the problem of oxygen diffusion, eventually even having an internal skeleton results in difficulties principally because, as Haldane pointed out, the strength of supporting leg bones grows as length squared (L2), whereas the weight of the body grows as length cubed (L3). The eventual result is disaster, the legs having to become ever larger, more massive and yet heavier in order to support the vast weight of the body. The exception is large sea animals, such as whales, in which the massive body weight is supported by the buoyancy of water rather than by the skeleton as such.
A video of the blood flowing through the heart, showing the spiral movement, and models of the action of a symmetric and an asymmetric heart can be seen at www.nature.com (Kilner et al., 2000).
![]() WWW☜ 5:8
WWW☜ 5:8
There are also other very exciting fossils coming out of China at present which will almost certainly help to clarify the origins of the vertebrates (Chen, Huang, & Li, 1999).
The literature on the evolution of vertebrates of exceedingly technical and very confusing. An excellent and relatively accessible account can be found in Gee (1996).
In writing this entire section I am uncomfortably aware that Dick Jefferies, like any good scientist, is continually revising his views on the basis of new evidence. Some of his published ideas are therefore no longer entirely his strict beliefs (although he does suggest that it would not be sensible for me to revise the text on the basis of as yet unpublished opinions). Here, in an e-mail of 16th November 2000, are Dick's current thoughts, for which I am exceedingly grateful:
"Your account of dexiothetism is fine except for a change of opinion on my part which you could not have known about. Namely, I have recently decided, on sequence evidence, that the correct basic cladogram for the deuterostomes is probably ((hemichordates + echinoderms) chordates) rather than (hemichordates (echinoderms + chordates)). This means that the group Dexiothetica (=echinoderms + chordates) is paraphyletic and ought to be forgotten. There are several implications from this:
1) I believe that dexiothetism certainly occurred, but that present-day hemichordates have lost almost all of the resulting asymmetries;
2) Cephalodiscus no longer seems to be very primitive and the bilaterally symmetrical "Cephalodiscus-like ancestor" is no longer closely related to Cephalodiscus - indeed, the sequence evidence implies that the latest common ancestor of extant hemichordates would have been a worm-like enteropneust, not a pterobranch resembling Cephalodiscus:
3) lurking among the fossils which I placed in the stem group of the echinoderms in a paper that I co-authored in 1996 (Jefferies, Brown & Daley. Acta Zoologica. 77, 101-122) must be members of the stem-group of the hemichordates as well as stem-group Ambulacraria (hemichordates + echinoderms) and true stem-group echinoderms - I think I know which fossils were the stem-group hemichordates but at present it is only a hunch:
4) the calcitic skeleton, restricted among extant forms to the echinoderms, has thus been lost four times among the deuterostomes, not three times as I formerly believed".
I am also aware that other researchers are reconsidering some parts of the theory (David et al., 2000). When deep experts disagree there is little the rest of us can do except sit and watch with fascination and await a new consensus. Until that arrives we can merely speculate on the implications of the ideas which are already floating around.
![]() WWW☜ 5:9
WWW☜ 5:9
This account is mainly based on that in Gee (1996 p.10). Recent molecular analyses suggests that a group called the Acoela are the most primitive Bilateria (Ruiz-Trillo et al., 1999).
![]() WWW☜ 5:10
WWW☜ 5:10
Dick Jefferies is undoubtedly a scholar’s idea of a scholar. I once met him at his office in the Natural History Museum, and spent several hours looking at specimens. At one point I asked him how many examples were known altogether of a particular species: “Oh, about half a dozen. They are all on the window sill over there!”. Dick is unashamedly a cladist in his interpretation of both living and fossil forms (Gee, 2000 pp.74-76), and approvingly cites Hennig’s dictum that “ ‘Das Kriterium der Wahrheit ist Vereinbarkeit’ – the criterion of truth is coherence (of results)” (Jefferies, 1986 p.15).
![]() WWW☜ 5:11
WWW☜ 5:11
Gee (1996 pp.201-286) provides a good critical account of the strengths and possible weaknesses of Jefferies’ theory. Even if it is not correct in all its details, and few scientific theories are ever correct in all their details, Jefferies’ theory is quite the best that we have at present, and it provides a serious and realistic attempt to create the sort of theory that is needed. Jefferies (1986 p.vii) particularly emphasises that it was Torsten Gislén who first proposed, seventy years earlier, that chordates arose from the calcichordates, but his ideas were almost totally ignored during his lifetime (Gislén, 1930).
Jefferies has commented about the reconstruction of the movements of Rhenocystis, this is
“palaeopsychology”, observing the recorded behaviour of organisms long, long dead. There is
something extremely moving
![]() about seeing the fossilised tracks or footprints of animals. One only
has to think of the human footprints at Laetoli (Hay & Leakey, 1982) or the recently discovered
dinosaur tracks in Greenland (Gatesy et al., 1999).
about seeing the fossilised tracks or footprints of animals. One only
has to think of the human footprints at Laetoli (Hay & Leakey, 1982) or the recently discovered
dinosaur tracks in Greenland (Gatesy et al., 1999).
![]() WWW☜ 5:12
WWW☜ 5:12
Gee (1996) states very clearly what our response should be to such ideas:
“...no idea, however odd, should ever fail simply because the reader finds it outrageous. Strangeness and novelty, after all, are attributes to be welcomed by science, for the oddball notion of today might be the orthodoxy of tomorrow. ... Even the most outré idea deserves serious consideration, and if it is found to fail, it will be by the standards of scientific investigation...”
Popularising science, writing interestingly, and also being accurate are extremely difficult things to reconcile. As a marvellously witty description of Jefferies’ theory which is wrong in almost every substantial respect I cannot resist quoting from an article in The Times of 1993:
“...one academic theory aired in recent years traces the starboard tendency to one of man’s less successful Jurassic ancestors, a sea-bed dwelling creature with a rudimentary left ear in its anus, causing it to lie permanently on its side in order to listen for the approach of predators and grab passing lemon sole with its right paw” (Hamilton, 1993).
There is some support for Jefferies' conjecture from work in developmental embryology, where there is a similarity between the cascade of signalling molecules which specify left and right and those which specify dorsal and ventral (Whitman & Mercola, 2001c).
![]() WWW☜ 5:13
WWW☜ 5:13
Richard Dawkins (1997 pp.204-210) considers the theoretical advantages of symmetry in more detail. That organisms do indeed tend to evolve symmetry when there is selection for speed is shown in an intriguing study in which robots were jointly selected both for the efficiency of their neural networks and for the body forms to which the networks were connected (Lipson & Pollack, 2000).
Well, dragons are mostly four limbed (and indeed in some from the 13th and 14th centuries they are four-limbed, “bipedal and winged and saurian in shape”, as it has been put (Hebron, 1985 p.3). However mythology (and the cover of Harry Potter and the Goblet of Fire) often shows dragons that have four legs and two wings, and that is not a vertebrate pattern, which is invariably tetrapod in its primitive form. The same problem has been pointed out with many conventional representations of angels, where there are two arms, two legs, and a pair of wings on the back, which does not fit the vertebrate pattern. But then, angels probably didn’t evolve either.
![]() WWW☜ 5:14
WWW☜ 5:14
This can be seen easily in any flatfish from a fishmonger. If it is held vertically upright then the basic body layout is the same as any other fish, with the exception that the eyes are in the wrong place, both being on one side. In particular the mouth can be seen now to have two lips, an upper and a lower, in an arrangement like that of any other fish.
The twisting of the flatfish's eye involves a number of re-arrangements, and in particular the nerves from the eyes to the brain, which normally cross at the topic chiasm, get an additional twist in them (Policansky, 1982).
Although the selective advantages of being a flatfish flat on the seabed seem clear enough, the mechanism by which flatfish attained this outcome is less than clear. In particular, how did the eye migration mechanism develop? And as with so many things in biology, surely it would have been a disaster to have the eye move only part of the way. So what were the intermediate states, and how were they advantageous?
![]() WWW☜ 5:15
WWW☜ 5:15
It is possible by transplanting tissue between embryos or applying beads to the embryo which been soaked in growth factor, to produce an additional limb which is the mirror-image of one already present on the same side, or to produce ‘mirror polydactyly’ in which there are extra fingers on an arm, with two thumbs in the middle, two index fingers beside them, and two middle fingers on the outside. See Larsen (1998 p.225) for illustrations of some of the defects. None of these cases, though, involve novel mechanisms as far as the mechanism of right and left determination is concerned.
It is intriguing that, rather like Conan Doyle’s dog that didn’t bark in the night, it is much easier in biology to ask what does occur and why, rather than to ask why certain problems do not arise and to ask why not. But each can be equally informative in theoretical terms. It is the absence of certain types of defect that is so instructive in this case.
![]() WWW☜ 5:16
WWW☜ 5:16
Although it sounds far more trivial, one of the problems of conditions such as cebocephaly is that, unlike cyclops in which the defects are so gross that the fetus usually dies, the less severely defects of cebocephaly are usually compatible with life. Larsen (1998 p.267) has a picture of a child with the problem. One dreads to imagine the stigmatisation suffered by such a child; it is reminiscent of the opening lines of Goffman’s Stigma, with its quotation from the novel, Miss Lonelyhearts, by Nathanael West: “...I would like to have boyfriends like the other girls and go out on Saturday nites, but no boy will take me because I was born without a nose – although I am a good dancer and have a nice shape and my father buys me pretty clothes” (Goffman, 1963). Although there are many causes for these rare syndromes, some genetic, one cause which is to a large extent avoidable is fetal alcohol syndrome due to excess maternal drinking.
Developmental biologists who discover new molecules are entitled to call them what they like. Although at the time when Sonic hedgehog was named it probably seemed clever and trendy, already it merely looks rather silly, and in a few decades it will be in as dead a language as the Latin and Greek names about which students presently complain. For a list of Drosophila genes, such as heartless, or dumb, or ken and barbie, along with their etymologies, see www.flynome.com.
‘Knockout mice’ (Chiang et al., 1996) in which particular genes of importance in development are targeted and removed have transformed our understanding of the role of particular genes in embryology. Newer techniques are also allowing genes to be knocked out just for short periods in development, so that effects later in development can be observed, or for genes to be knocked out and then degraded or altered versions 'knocked-in', to find out how which parts of the gene are of particular importance.
The establishment of the mid-line is in part controlled by slit (Harris & Holt, 1999). One effect of particular neurobiological interest is in directing nerves to their proper destination. In the brain it is essential that some nerve fibres cross over to the other side, whereas there are others that must stay on the side on which they start. And if fibres do cross the midline it is crucial that those same nerve fibres stay on that other side, rather than crossing back over the midline. The whole process is controlled by the product of a gene called slit (Harris & Holt, 1999).
Failure to establish a midline is probably also in part responsible for errors of lateralisation in conjoined twins (Burdine & Schier, 2000), in which monozygotic twins fail to separate properly.
![]() WWW☜ 5:18
WWW☜ 5:18
The names of the condition are almost as confused as the body parts, isomerism defect also being known also as Ivemark syndrome (Ivemark, 1955), situs ambiguus, visceral heterotaxy, laterality sequence, isomerism sequence, and asplenia/polysplenia syndrome (Burn, 1991). There are also the conditions known as the heterotaxias, in which some organs are reversed and others are not. This is not quite the same as isomerism but can be equally problematic, with the typical difficulty being in the connections of the heart, so that heart failure often results.
The causes of isomerism defect are not known, but there is evidence that left isomerism runs in families (Arnold, Bixler, & Girod, 1983) and is associated with diabetes in the mother (Splitt et al., 1999). That raises the interesting possibility that right and left isomerism have different underlying aetiologies.
In isomerism defect the problem is once again probably to do with the midline, keeping those things which are the left’s on the left and those things which are the right’s on the right. In fact when one looks carefully at individuals with isomerism defects they often also have developmental malformations of the midline itself (Goldstein, Ticho, & Fishman, 1998, Izraeli et al., 1999; Ticho, Goldstein, & Van Praagh, 2000). The same situation is also found in other animals, such as mice, zebrafish and the frog, Xenopus. The importance of the midline may not be merely in maintaining a physical midline barrier between the right and left halves but in actively suppressing right-expressing genes on the left and vice-versa (Izraeli et al, 1999 p.693). Analogies involving driving and motor cars seem to come naturally when trying to think about such problems, and a recent elegant one criticises an overly simplistic view of early embryos as being like “the tranquil lanes of a rural area”, and suggests they are more like “the tangled network of highways around Los Angeles”. And something is needed to control such traffic:
“If we look at ... the region either side of the midline, we see heavy traffic in all directions. Heart precursors migrate towards the midline; neural-crest cells migrate away from the dorsal neural tube; germline cells move towards the posterior gonadal ridge; and muscle precursors migrate from next to the midline towards the sides of the embryo to form slow muscle cells. Many [molecules] are required to direct this cellular traffic” (Driever, 2000 p.142).
It reminds me of those trunk roads which existed when I was a child. Instead of being like motorways, with several lanes in each direction, separated by a clear crash barrier to stop anything going from one side to the other, there were instead just three lanes, one in each direction, and a middle lane, officially called the ‘overtaking lane’, but known to everyone as the ‘suicide lane’ because of the disastrous crashes that occurred at regular intervals, throwing debris to left and right of the road. The absence of a clearly demarcated midline is not a good idea.
![]() WWW☜ 5:19
WWW☜ 5:19
Robert Burns, To a Mouse (1786).
For many paired organs there are actually slight directional asymmetries, consistent population level differences between the two sides as well, but I am ignoring these for present purposes. However in general the right arm is slightly longer than the left arm (Latimer & Lowrance, 1965; Ludwig, 1932 p.263), the left leg is slightly longer than the right leg (Latimer & Lowrance, 1965), and so on.
Although I have described fluctuating asymmetry in terms of the simple difference in size of the two sides, it is more accurate to calculate the relative difference, using some formula such as (R-L)/(R+L) to take differences in overall size into account. A child and an adult can then have the same relative difference despite having very different sizes overall. If there are consistent directional asymmetries, then the best measure of fluctuating asymmetry is the standard deviation of the difference score, after adjustment for overall size differences (Palmer & Strobeck, 1986). For superb reviews of the whole question of the measurement of fluctuating asymmetry and its subtleties, see Palmer and Strobeck (2001a) and Palmer (2000a).
![]() WWW☜ 5:20
WWW☜ 5:20
The development of the two sides, based on the instructions in the genes, is not like two guided missiles, aimed at the same target, and with error correcting software so that if either missile starts to move slightly off target then its trajectory will be altered to correct the problem. The cells forming the two incisors or the two arms or the two legs are instead like ballistic rockets: they are pointed in the right direction, given their instructions, launched and then there is nothing anybody can do to determine exactly where they land. Should a storm blow one of them around en route then the rocket ends up somewhere other than where it was intended. The question is not whether both rockets will hit the target but rather how far apart will they be when they land, for it is nigh on impossible that they should hit at precisely the same place.
Fluctuating asymmetry is ultimately due to chance alone. This form of chance must be contrasted with the sort of chance or random variation that is typically included in statistical analyses which is due either to measurement error or sampling variation, both of which can in principle be measured, accounted for and potentially eliminated. The chance variation in fluctuating asymmetry is a different sort, and in its final form it has been described as ‘deep chance’, a form of noise implicit in the Brownian motion of particles, and ultimately in the randomness of quantum fluctuations themselves.
Although the direction of asymmetry in individuals varies randomly from generation to generation, it fluctuates, it is also the case that fluctuating asymmetry in the sense of the extent of variance shown is probably heritable (MÆller & Swaddle, 1997 pp.113-119; Whitlock, 1996), although of course the direction of such variability is still not heritable. If your parents are very asymmetric then you are more likely to be asymmetric, but which way round the asymmetry will be is random.
I have presented a fairly conventional view of fluctuating asymmetry. There are however a number of problems in its interpretation, particularly as an indicator of genetic fitness; see amongst others, MØller & Pomiankowski, 1993; Bjorksten, Fowler, & Pomiankowski, 2002; David et al., 1998; Bjorksten et al., 2000; David et al., 1999.
![]() WWW☜ 5:21
WWW☜ 5:21
Genes often do not have the effect in different contexts. This is most obvious in the completely inbred mice which are so much used in laboratory research. All are identical to one another and often a gene can have a particular effect in them. However that same gene when introduced into another inbred strain will have a completely different effect. Geneticists refer to this as an effect of the ‘genetic background’.
On a technical note it must be emphasised again that instability, environmental disruptions, or whatever, do not alter the mean or the location of the distribution of left-right differences, which must stay at zero, but instead it increases the variance, and so more individuals will have larger right-left differences, although still precisely half will have right larger than left and half have left larger than right. In practice one also measures several or many characters at the same time and combines them statistically, and the individual characters can have different directions. That also allows the measurement of variation within the organism as well as between. It should also be emphasised that fluctuating asymmetry is a measure that applies to individual traits within an organism, whereas developmental stability applies to the organism as a whole, with important theoretical implications for the distribution of measures of asymmetry (Gangestad & Thornhill, 1999).
Although I have discussed the topic of fluctuating asymmetry / developmental instability as if it is well accepted by all those in the field, there are undoubtedly many theoretical problems associated with the area, some to do with measurement, and others to do with problems of replication (Palmer, 2000a).
![]() WWW☜ 5:23
WWW☜ 5:23
Sitwell continues, “But radiance is a better word to match her quality, I think, than beauty, and a certain sad radiance still clung to her”. There is a painting of Lady Sitwell as a girl in the original edition of Left hand, right hand! (Sitwell, 1945, and a rather poor profile photograph in The Sitwells (National Portrait Gallery, 1994 p.31), in which the characteristic Sitwell nose is just about visible.
Data on the relationship between fluctuating asymmetry and sexual selection and breeding success, as well as other biological parameters, can be found on the web-site which accompanies Møller and Swaddle (1997), www.oup.co.uk/MS-asymmetry/. There is some evidence that birds with slightly asymmetric wings and tail-feathers are less good at flying, although that of course may reflect aerodynamic problems themselves, rather than developmental stability as such (MÆller & Swaddle, 1997 pp.159-172).
For a marvellous semi-spoof on the whole issue of sexual selection and fluctuating asymmetry, see Palmer and Hammond (2000b).
![]() WWW☜ 5:24
WWW☜ 5:24
Embryology was even more difficult for the founders in its early, heroic stages. The survival of operated embryos was poor, and since there were no facilities for culturing organisms out of their natural season, amphibian embryos were only available for a short time between April and early July: “the pressure to continue operating throughout waking hours during the season cost the Nature-loving Spemann dearly, in foregoing spring over some forty years” (Horder & Weindling, 1986 p.193).
Embryologists use a number of terms to describe the fertilised egg of different species at different stages. For simplicity I have used embryo throughout.
![]() WWW☜ 5:25
WWW☜ 5:25
When in 1886-7 Hans Spemann had “a touch of tuberculosis” and spent the winter in a Swiss health resort, high in the mountains, a copy of Weismann’s later book (1892) was the only scientific work he took with him: “I found here a theory of heredity and development elaborated with uncommon perspicacity to its ultimate consequences. ... This stimulated experimental work of my own” (Hamburger, 1988 p.9). In thinking about Weismann and Roux it is important to remember that at thetime no one had the faintest idea what the genetic material might be, and how it might be organised. Neither had Mendelian genetics yet been rediscovered.
Viktor Hamburger, who perhaps should have received the Nobel Prize for experimental embryology in 1986, and who was publishing on the history of the subject almost until his death, died on 12th June 2001, just short of his 101st birthday.
Reflection suggests that things could not possibly be as simple as the mosaic theory suggested. Although always couched in terms of right and left halves, in the absence of anything defining front-back and top-bottom, then right and left could not even be determined. In fact Roux did also find embryos in which the back or the front of the embryo was missing, although textbooks tend to concentrate on the left-right situation.
According to Spemann, the problem of Roux's experiment seems to be that although one of the cells was no longer alive, “effects of a more simple and purely mechanical kind might have issued from it” (p.21). Spemann actually uses the term “vital” rather than alive, and he puts it in quotations. Only when his autobiography was published posthumously in 1943 did it become apparent that Spemann was a vitalist, believing that “all parts of an organism possess an element of consciousness” (Hamburger, 1999 p.242). Driesch’s experiment used the sea urchin, which is an echinoderm and therefore one of the most closely related of all the living invertebrates to the chordates. Subsequent experiments showed that the same result was found in frogs. It was also shown that if one used Roux’s hot-needle technique and then removed the dead cell then the organism developed normally (Spemann, 1938 pp.24-25).
The result of Roux's experiment was not really explained until many years later, when the Nieuwkoop centre was identified, and it became apparent that the plane of cleavage had already passed through it, and that the remaining live cell did not ‘know’ that the other cell was dead (Wolpert et al., 1998 p. 67).
![]() WWW☜ 5:26
WWW☜ 5:26
Spemann received the Nobel Prize in 1935, three years before Hitler instituted national prizes for the arts and sciences and banned Germans from accepting the Nobel Prize (Horder & Weindling, 1986). The Prize was awarded for the discovery of the ‘organizer’, a region of the early embryo which acted as a sort of ‘master switch’ for development. Transplantation of the organizer from one embryo to another would result in the development of two head regions. Although the crucial experiment was carried out with Hilde Mangold, the prize was awarded to Spemann alone since Mangold, who was then 26 years old, had died of burns in September 1924 after a gasoline heater exploded in her kitchen (Hamburger, 1988 p.173). There have recently been suggestions that the key organiser experiment had also been performed earlier, in 1909, by an American zoologist, Ethel Browne, and that Spemann was aware of those results (Lenhoff, 1991). The name of the organiser has itself been controversial, and Spemann himself referred to it on occasions as the ‘leader’ or the ‘Führer’, and he seems to have used the term in a metaphorical sense in a political context (Horder & Weindling, 1986).
Reviewing these and other experiments a few years later Huxley and de Beer (1934 pp.75-77) also note how double-headed monsters also occur spontaneously in trout, both in the wild and in hatcheries, and that the right hand member shows situs inversus on about half of occasions. Over the years the explanations of the difference between the right and the left embryo have been many and varied. As Oppenheimer (1974pp. 871-2) says, “The interpretations have varied according to their times: asymmetrical microstructure, intrinsic bending tendency, material deficiency, asymmetric induction, localised asymmetry factor, left dominance in a left-right physiological gradient, physiological determination, reversal by regulatory movements, reversal as a result of developmental arrest, and many others have been called upon as explanatory”.
![]() WWW☜ 5:27
WWW☜ 5:27
![]()
The British Association meeting in Toronto in 1897 was the second of its meetings to be held abroad; the first one in 1884 in Montreal was more of a social than a scientific success, being described as “the longest and most lavish ‘picnic’ in the Association’s history” (Worboys, 1981 p.175).
Tantamount seems to be an amalgam of several figures. Naomi Mitchison (1965 p.53), the novelist and sister of JBS Haldane, the physiologist, claimed that Tantamount was based on her father, John Scott Haldane, himself a physiologist who was notorious for his eccentricities. The opportunity would have been there, as Aldous Huxley and the Haldanes were close friends. The parallels are also fairly close, Haldane having a private laboratory in his own house, working in his laboratories to all hours, seeming to be as unworldly as it is possible to be, and in later life becoming more interested in the non-material, spiritual aspects of science (Clark, 1968). However Haldane himself disliked experimenting on animals, preferring to experiment on himself (and his children). Furthermore Haldane was never interested in embryology. Onto Haldane’s personality seems therefore to have been grafted Julian Huxley’s own experimental interests and methods, as Deery (1996 p.34) seems to imply (although JBS Haldane seems to have been confused with JS Haldane). Tantamount also has overtones of the heterodox ideas of Bastian 1913, who rejected Pasteur’s experiments on spontaneous generation, work Aldous Huxley may well have been aware of, his grandfather, Thomas Henry, attacking it in 1870 (Bastian, 1913 p.8), and Aldous does cite Bastian’s other work in Point Counter Point. Finally, there is a strong hint of Sir Michael Foster (1836-1907), the Cambridge physiologist, who gave the Presidential Address to the Physiology section at the meeting in Toronto in 1897. He mentioned Du Bois Reymond, who had died since the previous Canadian meeting, in a rather barbed comment: “who, if he laboured in a narrow path, set a brilliant example of the way in which exact physical analysis may be applied to the physiology of living beings, and in other ways had a powerful influence of the progress of physiology", and he also discussed the development of “the new physiological chemistry” (which would have included the role of osmosis), and singled out the work of Emil Fischer who was at that time sorting out the absolute structure of chiral molecules. Finally, in a remarkable last passage, referring in part to the anti-vivisectionists, and reminiscent of the scientific evangelism of Tantamount, he described how, “We physiologists are sorely tempted towards self-righteousness, for we enjoy that blessedness which comes when men revile you and persecute you and say all manner of evil against you falsely". It seems difficult to believe that Aldous Huxley had not browsed through the proceedings of the Toronto meeting, and did not know Foster, who gained the Chair of Physiology at Cambridge on the basis of T H Huxley’s advice (Geison, 1978).
![]() WWW☜ 5:28
WWW☜ 5:28
In the year he died, Aldous Huxley, in response to CP Snow’s The two cultures, published a little book about the relations of science and literate which he contrasted in terms of pairs of opposites: “Public and private. Objective and subjective. The world of concepts and the multitudinous abyss of immediate experience. The simplified, jargonized purity of scientific discourse and the magical, many-meaninged purity of literature” (Huxley, 1963 p.36). One is crying out to be on the right and one on the left... Huxley worries about the paradox that “the poetry of this most scientific of centuries should be ... less concerned with science than was the poetry of times in which science was relatively unimportant...”. In part it is, he says, because “‘Popular science’ is a new art form, partaking simultaneously of the text book and the reportage, the philosophical essay and the sociological forecast”.
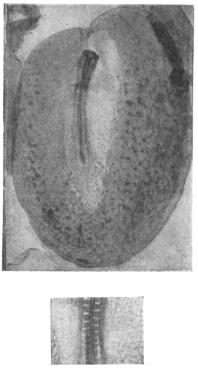
The elements of experimental embryology (Huxley & de Beer, 1934) has been described as “coming at the high point of optimism with regard to the prospects for a chemical identification of the organiser” (Horder & Weindling, 1986 p.226).
Art was clearly imitating Life rather directly in the experiments Huxley described in Point Counter Point, because Huxley and de Beer’s book has a whole section devoted to the question of how temperature gradients affect the development of embryos (Tazelaar, 1929,Tazelaar, Huxley, & De Beer, 1930). As well as descriptions of experiments on newts and frogs, there is a photograph of a chick embryo, published originally in 1928, the same year as Point Counter Point, in which the higher temperature on the left hand side has not only meant that the whole embryo is curved, but also on that side there are more somites, the precursors of the vertebrae which make up the backbone (figure 14). Although such experiments were technically ingenious, and though they did occasionally seem to shift the position of the midline of the organism (Huxley & de Beer, 1934 p.342), ultimately they did little to resolve the problem of why the heart was usually on the left and occasionally was on the right.
![]() WWW☜ 5:29
WWW☜ 5:29
There were of course exceptions, and some useful, solid work did occur. Oppenheimer (1974) reviews the work in particular of von Woellwarth (“a sort of intellectual grandson of Spemann”), and von Kraft (who has recently published in English an overview of three decades of studies – see Von Kraft, 1999). See also Wehrmaker (1969).
The key theoretical papers which influenced both psychologists and embryologists were by Michael Morgan, one of my PhD supervisors, who is now at City University in London (and who is the winner of the 2001 Wellcome Trust Prize to write a popular science book) , and Michael Corballis who is at the University of Auckland in New Zealand. The 1978 papers (Corballis & Morgan, 1978, Morgan & Corballis, 1978) referred to Layton’s work, and the reply to commentaries (Morgan, 1978) also mentioned Afzelius’s study. The papers of Morgan and Corballis also mention, amongst others, the work of Wilhelmi, Harrison and Von Woellwarth.
![]() WWW☜ 5:30
WWW☜ 5:30
There had also been an earlier report of a similar mutation causing situs inversus but the strain was weak and died out (Tihen, Charles, & Sippel, 1948).
Although Layton (1976) refers to the defect as being “situs inversus viscerum”, there is a suggestion that many of these animals do not have pure situs inversus, but a high proportion have some heterotaxia, with some organs being mirror-reversed and others being in their normal location (Brown & Wolpert, 1990). Layton himself reported some heterotaxias of the heart and of the venous system in a subsequent paper (Layton, 1978). In this respect the mice may be different from the majority of humans with situs inversus who do not show heterotaxia.
In the twelve years since the original paper was published (Layton, 1976), the mice had gone through 32 generations, equivalent to about eight or nine centuries of human breeding. One can see why the mouse is such a favourite animal for genetic research.
It is also possible to tell in utero whether a mouse embryo has situs inversus since a normal mice coils in a right-handed direction, whereas a mouse with situs inversus has a left-hand coil (Layton, 1976, Figure 1).
At a single genetic locus, such as iv, an individual will have two copies of the gene, one from each parent. The normal gene (allele), which is called the ‘wild’ type and occurs in the common field or house mouse, is conventionally shown as +. If a mouse has received the same gene from each of its parents, either two + or two iv genes, it is said to be homozygous, and its genotype is either +/+ or iv/iv. If instead it has received different genes from the two parents, it is said to be heterozygous and has a genotype +/iv. The only mice which show situs inversus are those with the iv/iv genotype.
An important theoretical possibility that had to be ruled out in explaining the 50% incidence of situs inversus in the homozygous iv/iv mouse was that the gene had what is called partial penetrance, meaning that other factors were also important in determining on which side the heart would be placed. That explanation was however excluded by Layton taking mice which were all iv/iv and interbreeding them (so all their offspring had also to be iv/iv). He then looked at whether the proportion of offspring with situs inversus related to whether or not the parents had situs inversus. It did not. Whether both parents had their heart on the left, both had their heart on the right, or one had the one on one side and one on the other, in each case exactly a half of the offspring had their heart on the right and a half on the left.
![]() WWW☜ 5:31
WWW☜ 5:31
Although the syndrome is known as Kartagener’s syndrome after the Swiss paediatrician who described four cases in his 1933 paper (Kartagener, 1933), Kartagener was not actually the first person to describe the condition, Siewert describing it in 1904 (Siewert, 1904). History seems to have had its revenge on Kartagener because there seems to be utter confusion as to how to describe his name, researchers varying between Kar-ta-gay-ner and Kar-taj-en-er. And in recent years, ‘primary ciliary dyskinesia’ has more frequently been used (Parraudeau et al., 1994).
It is not clear from this particular patient's case history whether the patient himself was aware of his situs inversus, and although it is implied that he was infertile, it is not stated whether he had any children or had tried unsuccessfully to have any. That he continued to smoke at such a level with such symptoms suggests this was the first time he had been diagnosed as having Kartagener’s syndrome (Parraudeau et al, 1994). Although it is unusual for the condition to be diagnosed in mid-life, there is a report in the literature of a patient not diagnosed until the age of 75 (Gomez-de-Terreros-Caro et al., 1999).
Not all male patients with Kartagener’s syndrome are infertile, and in some cases the sperm do seem to have some mobility, albeit not normal. It is for that reason the syndrome is now called primary ciliary dyskinesia, meaning impaired movement, rather than immotile cilia syndrome, which implies a complete lack of movement. These patients are probably also fertile if intracytoplasmic sperm injection techniques are used for getting the sperm into the egg.
![]() WWW☜ 5:33
WWW☜ 5:33
In a review nearly a quarter of a century later, Afzelius (1999b) quotes his earlier view that the cilia “have a certain position and a fixed beat direction and that their beating somehow is instrumental in determining the visceral situs”. He acknowledges though that a crucial problem was that normal 9+2 cilia beat back and forth in a whip-like motion, and hence there would still have to be something that orientated them to left and right of the organism.. The same theoretical problem does not occur with the inherently chiral monocilia.
![]() WWW☜ 5:34
WWW☜ 5:34
The drug acetazolamide is a carbonic anhydrase inhibitor, and is used among other things for the treatment of glaucoma, as a weak diuretic and an anti-convulsant, and for preventing acute mountain sickness.
Brown and Wolpert's key theoretical paper began, “The development of handed asymmetry is a deep and neglected problem” (Brown & Wolpert, 1990).
In their theoretical model, Brown and Wolpert wrote, in a phrase that might be seen as prescient: “...we propose an interaction between the oriented F molecule and the polarity of the cells with respect to the midline. For example, if the F molecule caused transport of some other molecules in the direction of its ‘arms’ ... then the transport would be towards the midline on the left side and away from it on the right side” (Brown & Wolpert, 1990 p.3).
There have been two important integrative meetings subsequent to the Ciba meeting in 1991, one in the Banbury Centre at Cold Spring Harbor in November 1997, and then at the Juan March Foundation in Madrid in April 2001 (Wright, 2001).
![]() WWW☜ 5:35
WWW☜ 5:35
There is an inevitable problem in writing about left and right, and in particular of presenting pictures
of left and right; does one place the left of an object to the left (for verbal consistency) or to the right
(as it looks when one faces it, just as when we look face to face at another person their left hand is
on our right). Biologists have not been consistent
![]() , and that is reflected in the diagrams and pictures
in this chapter. Partly it reflects the facts that dissections are usually carried out from the ventral
rather than the dorsal surface, so that left and right seem to be reversed
, and that is reflected in the diagrams and pictures
in this chapter. Partly it reflects the facts that dissections are usually carried out from the ventral
rather than the dorsal surface, so that left and right seem to be reversed
![]() . And likewise microscope
slides produced on a microtome are usually, in effect, reversed because of the way the blocks are
sliced. However many microscope images are nowadays digitised, and so it is easy to 'flip' the
image so that left and right are to the left and right of the viewer. Although I considered being
consistent in this chapter, in practice there seemed to be no easy way to do it sensibly, and I have
therefore left the images as they were presented in the originals.
. And likewise microscope
slides produced on a microtome are usually, in effect, reversed because of the way the blocks are
sliced. However many microscope images are nowadays digitised, and so it is easy to 'flip' the
image so that left and right are to the left and right of the viewer. Although I considered being
consistent in this chapter, in practice there seemed to be no easy way to do it sensibly, and I have
therefore left the images as they were presented in the originals.
![]() WWW☜ 5:36
WWW☜ 5:36
Although the bulging of the heart tube is the most visible sign of the asymmetry of the developing heart, there is a slight asymmetry of Hensen’s node visible before the heart tube is present (Cooke, 1995).
For a superb review of the various signalling molecules that have been implicated in left-right asymmetry, see Mercola and Levin (2001b).
![]() WWW☜ 5:37
WWW☜ 5:37
The classic example in biology of correlation not proving causation is that in the countries of northern Europe there is a high correlation between the numbers of storks and the birth rate. This does not however prove that the babies are brought by the storks, but merely shows that the times of year when births are highest and the times of year when storks migrate are themselves similar.
Although the story as presented about Sonic hedgehog seems straightforward, it has since become very complex, not least because Sonic hedgehog seems to behave differently in different species; for instance mice with Sonic hedgehog knocked out do not seem to have an increased rate of situs inversus. The explanation of these problems probably involves homologous cascades in different organisms, with a wide range of intermediary substances (Esteban et al., 1999, Capdevila et al., 2000, Levin et al., 1997, Ryan et al., 1998).
The number of molecules involved in signalling left and right seems to increase every day. In September 2000 Michael Levin’s web-site described 24 genes that are asymmetrically expressed in embryos, and a further seven that although expressed symmetrically are involved in left-right asymmetry (134.174.168.95/ ~mlevin/lr_asymm.shtml). Whitman and Mercola (2001c) have since provided a useful diagram comparing the processes in chick, mouse, Xenopus and zebrafish. Why there should be such variability between species in what would one would have thought to be a heavily conserved process is still a mystery. However it is an accepted principle in developmental biology that the early stages of development are often very variable, being particularly dependent on the ways and places in which fertilisation occurs, and, in mammals, on the geometry and nature of implantation. Subsequent stages during early organogenesis are then much more similar ('conserved'). In laterality this is known as the 'left-right phylotypic stage' (Yost, 2001), and seems to focus on the roles of the nodal-related proteins and Pitx2. Later organogenesis can then once more become much more variable as a result of the different organisations and forms required by different species.
![]() WWW☜ 5:38
WWW☜ 5:38
![]()
In a subsequent paper it was found that left-right dynein was expressed in the node, just where Sonic hedgehog and similar substances would later be found to be expressed asymmetrically. But the problem was that it was present in equal amounts on the left and the right sides (Supp et al., 1999). The analysis of the iv mouse has been much improved by analysing the lrd mouse which has the gene for left-right dynein knocked out specifically, and unlike the iv/iv mouse this produces normal litter sizes and is generally more robust. A variant of the lrd mouse has also been produced which has, amongst other things, a neomycin gene inserted into the region near the gene. This had the entirely unexpected effect of producing about 70% of mice with situs inversus, thereby parallelling the reversal found in the inv mouse. One possibility, still to be investigated, is that the cilia in the lrd-neo mice also beat less efficiently, slowing transport across the node.
![]() WWW☜ 5:39
WWW☜ 5:39
A video of the beating monocilia can be seen at http://www.molecule.org/ cgi/content/full/4/4/459. The original paper by Nonaka et al (Nonaka et al., 1998) erroneously described the rotation as anti-clockwise, the error arising due to the high speed of rotation of the cilia and aliasing on a relatively low speed video recorder (see subsequent Erratum in Cell).
It had previously been generally thought that monocilia were immotile and mainly had a sensory role, providing a bridge between the environment of the cell, and the inside (e.g. Cohen & Meininger, 1987). Monocilia or primary cilia also occur widely, for reasons unknown, in any cell in culture which is provided with a limited food supply and becomes quiescent.
Embryologists, with the exception of Afzelius, had generally ignored the possibility that cilia might have an important role to play in directing development. One exception was Twitty (1928), who carried out experiments in the laboratory of Ros Harrison on transplanting cilia. In his intellectual autobiography he suggests:
“Altogether the ciliary project was a rewarding and entertaining initiation into experimental biology. I suspect that embryologists will find it profitable to return to these organelles when methods become available for effectively attacking phenomena of cellular polarity at the molecular level”. (Twitty, 1966 p.19).
![]() WWW☜ 5:40
WWW☜ 5:40
Brueckner and her team at first could not replicate the finding that the normal monocilia rotated, and only succeeded when they found exactly the right microscope: “They’re there. It’s real”, she eventually said Ainsworth, 2000. A video of normal and iv cells can be found at http://genome.chmcc.org/cilia/ Supp, Potter, & Brueckner, 2000. At a meeting in June 2001 Hirokawa described his iconoclastic finding of the beating monocilia and showed a slide with a picture of Galileo and his famous quotation, "But still it moves..."...
![]() WWW☜ 5:41
WWW☜ 5:41
The inv mouse was presented at a meeting at Cold Spring Harbor in 1992, and the paper was published the next year (Yokoyama et al., 1993), when Nigel Brown and Tony Lander described it as “one of those rare and remarkable papers announcing the completely unexpected” (Brown & Lander, 1993).The gene, also called the inversin gene, has been cloned (Mochizuki et al., 1998), and has been found to produce disruption of hepatobiliary system, which may account for the known association of situs inversus with biliary atresia (Mazziotti et al., 1999).
Although iv and inv look very similar when printed, inv is pronounced like the first syllable of ‘inversion’, whereas iv is pronounced as “eye-vee”.
The idea that the proportion of anomalous types cannot rise above 50% is related to a concept, which I have called Morgan’s principle (McManus, 1979, McManus, 1984, McManus, 1985). Based on the papers of Michael Morgan (Morgan, 1977, Corballis & Morgan, 1978, Morgan & Corballis, 1978), it says that paired alleles cannot produce enantiomorphic phenotypes – “genetic models of asymmetry should be asymmetrical”, as Morgan (1978) puts it. It is therefore highly unlikely that if one allele makes something be on the left then the other will make it produce the mirror image phenotype on the right.
![]() WWW☜ 5:42
WWW☜ 5:42
It was not completely impossible that the inv monocilia rotated in the opposite direction since it had long been known that bacterial flagellae, based on the protein flagellin, are capable of rotating both clockwise and anti-clockwise. A tiny change in the conformation of the protein alters the flagella from a right-handed to a left-handed helix (Berg, 1991; Samatey et al., 2001).
The flow in the inv mouse is clearly abnormal. Normally a bead starting at the right hand side
would reach the midline within about three seconds and get to the left edge within five seconds. The
movement in the inv mouse is not nearly so good, and half the beads had not reached the left edge
after ten seconds. There was also a suggestion of turbulence in the node with beads circling around,
rather than making their normal almost straight line path to the other side. Why though should the
flow be turbulent and slow? In fact there is another question lurking here which some readers may
already have worried about. Why even in the normal mouse should a rotating motion of the
propellers send the beads from right to left and not vice versa. After all if you stir the water in your
bath on one side it goes one way, but it comes back down the other side the other way. How can
the flow go consistently in only one direction? Hirokawa suggested that the one way traffic took
place not only because of the action of the rotating monocilia. What was important was that the
node is not a simple circular depression, but is actually shaped like a triangle pointing towards the
front of the organism. Although on their own or in a circular depression the cilia would produce
equal rightward and leftward flows, the effect of being in a triangular pit is to mean that the
rightward flow pass forwards, towards the top of the triangle, where its force would tend to be
dissipated
![]() . The result is then a leftward flow overall. Because the node is smaller and has a rather
longer, thinner shape in the inv mouse, the normal leftward flow is disrupted and slows down.
. The result is then a leftward flow overall. Because the node is smaller and has a rather
longer, thinner shape in the inv mouse, the normal leftward flow is disrupted and slows down.
As well as the abnormal nodal shape, it has also been suggested that the phase relationship between the turning of the inv monocilia may be disrupted, although it is very difficult to tell because they are turning so fast and they are so difficult to see easily (Okada et al., 1999 p. 462).
![]() WWW☜ 5:43
WWW☜ 5:43
Okada et al actually propose several models, but I concentrate here entirely on what they call the ‘delayed activation’ model (Okada et al, 1999 p.466).
Although it may sound a bit ad hoc for the molecule firstly to be activated and then to be deactivated, such processes are completely standard in biology. Many hormones are stored and transported as inactive pro-hormones, and only start working when something is snipped off one end. Likewise almost immediately after many neurotransmitters are released, enzymes start mopping them up in order to ‘turn off’ the response. Okada et al (1999 p.467) point out that Sonic hedgehog autocatalyses its own activation (Lee et al., 1994).
![]() WWW☜ 5:44
WWW☜ 5:44
I have enormous admiration for the work of Hirokawa's team. Having followed this confused
literature for thirty years and having been unable to see how the system could ever be made sense
of in terms of biology, Hirokawa’s work undoubtedly has the breath of genius. Not only does it
provide plausible explanations of almost all of the major phenomena that required explanation, but it
also feels like real biology. So much of what is nowadays called biological research is actually
nothing but manipulating strings of DNA base pairs within computers. The key thing that Hirokawa
and his colleagues did was actually to look down the microscope very, very carefully, and reconcile
what they saw with the sophisticated biochemical and genetic techniques they were also using. Like
all good research the nodal flow theory raises many questions of detail, and no doubt other
researchers will dot the is and cross a few ts, but the basic formulation seems unlikely to be
disproven
![]() . And even if it should prove to be wrong, it has raised more interesting questions and
looked in new ways at what were thought to be well understood processes than could have
imagined even five years ago.
. And even if it should prove to be wrong, it has raised more interesting questions and
looked in new ways at what were thought to be well understood processes than could have
imagined even five years ago.
A tricky problem for the nodal theory remains the difficult question of whether nodal flow is important in species other than the mouse, and there much confusion still reigns (Burdine & Schier, 2000), and other mechanisms have been proposed, such as the role of gap junctions in Xenopus (Levin & Mercola, 1998). It has also been suggested that the principle role of nodal cilia is not to generate asymmetry de novo, but instead is to amplify a pre-existing asymmetry is some species (Wagner & Yost, 2000). For the purposes of the present exposition I am not particularly worried as to whether the mechanism is the same in all species, as I am impressed by the remarkable finding that an entirely different mechanism from any that had been previously conceived could be responsible, even if only in one species. That alone makes the whole approach worthwhile, and it provides a beautiful story of how science progresses.
It would however be nice to have some more experimental demonstration that nodal flow was indeed important. Afzelius (Afzelius, 1999b) has suggested raising embryos in a viscous solution, to reduce the flows, and Wolpert has suggested using a jet of water to disrupt the flows (Ainsworth, 2000). Perhaps such experiments will have been done by the time this book is published, although they will be technically difficult.
The whole topic of molecular motors has been given new impetus recently by synthetic molecular motors being created chemically. Because the molecules of which they are made are chiral they rotate in a particular direction (Davis, 1999). Indeed, if the molecules were achiral then the motor would probably not be able to rotate at all.
Afzelius, B. A. 1999b, "Asymmetry of cilia and of mice and men", Int.J Dev.Biol., 43, 283-286.
Ainsworth, C. 2000, "Left right and wrong", New Scientist, 17th June, 40-45.
Altmann, S. L. 1992, Icons and symmetries, Oxford: Clarendon Press.
Arnold, G. L., Bixler, D., & Girod, D. 1983, "Probable autosomal recessive inheritance of polysplenia, situs inversus and cardiac defects in an Amish family", American Journal of Medical Genetics, 16, 35-42.
Bastian, H. C. 1913, The origin of life: being an account of experiments with certain superheated saline solutions in hermetically sealed vessels, 2 edn, London: Watts.
Berg, H. C. 1991, "Bacterial motility: handedness and symmetry," pp. 58-72 in Biological asymmetry and handedness (Ciba foundation symposium 162), G. R. Bock & J. Marsh, eds., Chichester: Wiley.
Bjorksten, T., David, P., Pomiankowski, A., & Fowler, K. 2000, "Fluctuating asymmetry of sexual and nonsexual traits in stalk-eyed flies: a poor indicator of developmental stress and genetic quantity", Journal of Evolutionary Biology, 13, 89-97.
Bjorksten, T., Fowler, K., & Pomiankowski, A. 2002, "What does sexual trait FA tell us about stress?", TREE (Trends in Ecology and Evolution), 15, 163-166.
Brown, N. A. & Lander, A. 1993, "On the other hand ...", Nature, 363, 303-304.
Brown, N. A. & Wolpert, L. 1990, "The development of handedness in left/right asymmetry", Development, 109, 1-9.
Burdine, R. D. & Schier, A. F. 2000, "Conserved and divergent mechanisms in left-right axis formation", Genes and Development, 14, 763-776.
Burn, J. 1991, "Disturbance of morphological laterality in humans," pp. 282-299 in Biological asymmetry and handedness (Ciba foundation symposium 162), G. R. Bock & J. Marsh, eds., Chichester: Wiley.
Capdevila, J., Vogan, K. J., Tabin, C. J., & Belmonte, J. C. I. 2000, "Mechanisms of left-right determination in vertebrates", Cell, 101, 9-21.
Chen, J.-Y., Huang, D.-Y., & Li, C.-W. 1999, "An early Cambrian craniate-like chordate", Nature, 402, 518-522.
Chiang, C., Litingtung, Y., Lee, E., Young, K. E., Corden, J. L., Westphal, H., & Beachy, P. A. 1996, "Cyclopia and defective axial patterning mice lacking Sonic hedgehog gene function", Nature, 383, 407-413.
Clark, R. 1968, J.B.S.: The life and work of JBS Haldane, London: Hodder and Stoughton.
Cohen, E. & Meininger, V. 1987, "Ultrastructural analysis of primary cilium in the embryonic tissue of mouse", International Journal of Developmental Neuroscience, 5, 43-51.
Cooke, J. 1995, "Vertebrate embryo handedness", Nature, 374, 681-681.
Corballis, M. C. & Morgan, M. J. 1978, "On the biological basis of human laterality: I. Evidence for a maturational left-right gradient", Behavioral and Brain Sciences, 2, 261-269.
David, B., Lefebvre, B., Mooi, R., & Parsley, R. 2000, "Are homalozoans echinoderms? An answer from the extraxial-axial theory", Paleobiology, 26, 529-555.
David, P., Hingle, A., Fowler, K., & Pomiankowski, A. 1999, "Measurement bias and fluctuatung asymmetry estimates", Animal Behaviour, 57, 251-253.
David, P., Hingle, A., Greig, D., Rutherford, A., Pomiankowski, A., & Fowler, K. 1998, "Male sexual ornament size but not asymmetry reflects condition in stalk-eyed flies", Proceedings of the Royal Society of London, Series B, 265, 2211-2216.
Davis, A. P. 1999, "Synthetic molecular motors", Nature, 401, 120-121.
Dawkins, R. 1997, Climbing Mount Improbable, London: Penguin Books.
Deery, J. 1996, Aldous Huxley and the mysticism of science, Basingstoke: Macmillan.
Driever, W. 2000, "Bringing two hearts together", Nature, 406, 141-142.
Elliot Smith, G. 1919, The evolution of the dragon, Manchester: Manchester University Press.
Esteban, C. R., Capdevila, J., Economides, A. N., Pascual, J., Ortiz, A., & Belmonte, J. C. I. 1999, "The novel Cer-like protein Caronte mediates the establishment of embryonic left-right asymmetry", Nature, 401, 243-251.
Gangestad, S. W. & Thornhill, R. 1999, "Individual differences in the developmental precision and fluctuating asymmetry: a model and its implication", Journal of Evolutionary Biology, 12, 402-416.
Gatesy, S. M., Middleton, K. M., Jenkins, F. A., & Shubin, N. H. 1999, "Three-dimensional preservation of foot movements in Triassic theropod dinosaurs", Nature, 399, 141-144.
Gee, H. 1996, Before the backbone: Views on the origins of the vertebrates, London: Chapman and Hall.
Gee, H. 2000, Deep time: Cladistics, the revolution in evolution, London: Fourth Estate.
Geison, G. L. 1978, Michael Foster and the Cambridge school of physiology, Princeton, NJ: Princeton University Press.
Gislén, T. 1930, "Affinities between the Echonodermata, Enteropneusta and Chordonia", Zool.Bidr.Uppsala, 12, 199-304.
Goffman, E. 1963, Stigma: Notes on the management of spoiled identity, Englewood Cliffs, NJ: Prentice-Hall.
Goldstein, A. M., Ticho, B. S., & Fishman, M. C. 1998, "Patterning the heart's left-right axis: from zebrafish to man", Dev.Genet., 22, 278-287.
Gomez-de-Terreros-Caro, F. J., Gomez-Stern, A. C., Alvarez-Sala, W. R., Prados, S. C., Garcia, R. F., & Villamor, L. J. 1999, "[Kartagener's syndrome. Diagnosis in a 75 year-old woman]", Arch Bronconeumol., 35, 242-244.
Hamburger, V. 1988, The heritage of experimental embryology: Hans Spemann and the organizer, New York: Oxford University Press.
Hamburger, V. 1999, "Hans Spemann on vitalism in biology: Translation of a portion of Spemann's Autobiography", Journal of the History of Biology, 32, 231-243.
Hamilton, A. 1993, "Cuddy-wifties right wronged sinistrals", Times, 14th August, 3-3.
Harris, W. A. & Holt, C. E. 1999, "Slit, the midline repellent", Nature, 398, 462-463.
Hay, R. L. & Leakey, M. D. 1982, "The fossil footprints of Laetoli", Scientific American, 246, 50-57.
Heaney, S. 1999, Beowulf, London: Faber and Faber.
Hebron, M. E. 1985, Statistical studies of the iconography of the dragon in Biblical texts of the 13th and 14th centuries, London: Chameleon Press.
Horder, T. J. & Weindling, P. J. 1986, "Hans Spemann and the organiser," pp. 183-242 in A history of embryology, T. J. Horder, J. A. Witkowski, & C. C. Wylie, eds., Cambridge: Cambridge University Press.
Huxley, A. 1963, Literature and science, London: Chatto &Windus.
Huxley, J. S. & de Beer, G. R. 1934, The elements of experimental embryology, Cambridge: Cambridge University Press.
Huxley, J. S. & de Beer, G. R. 1934, "The origin of polarity, symmetry, and asymmetry," pp. 60-82 in The elements of experimental embryology, Cambridge: Cambridge University Press.
Ivemark, B. L. 1955, "Implications of agenesis of the spleen on the pathogenesis of cono-truncus anomalies in childhood: analysis of the heart malformations in splenic agenesis syndrome, with fourteen new cases.", Acta Paediatrica, 44 (Suppl 104), 1-110.
Izraeli, S., Lowe, L. A., Bertness, V. L., Good, D. J., Dorward, D. W., Kirsch, I. R., & Kuehn, M. R. 1999, "The SIL gene is required for mouse embryonic axial development and left-right specification", Nature, 399, 691-694.
Jefferies, R. P. S. 1986, The ancestry of the vertebrates, London: British Museum (Natural History).
Kartagener, M. 1933, "Zur Pathogenese der Bronchiektasen. I Mitteilung Bronchiektasien bei situs viscerum inversus", Beiträge zur Klinik und Erforschung der Tuberkulose und der Lungenkrankenheiten, 83, 489-501.
Kilner, P. J., Yang, G.-Z., Wilkes, A. J., Mohiaddin, R. H., Firmin, D. N., & Yacoub, M. H. 2000, "Asymmetric redirection of flow through the heart", Nature, 404, 759-761.
Larsen, W. J. 1998, Essentials of human embryology, New York: Churchill Livingstone.
Latimer, H. B. & Lowrance, E. W. 1965, "Bilateral asymmetry in weight and in length of human bones", Anatomical Record, 152, 217-224.
Layton, W. M. 1976, "Random determination of a developmental process", Journal of Heredity, 67, 336-338.
Layton, W. M. 1978, "Heart malformations in mice homozygous for a gene causing situs inversus", Birth Defects, 14, 277-293.
Lee, J. J., Ekker, S. C., Von Kessler, D. P., Porter, J. A., Sun, B. I., & Beachy, P. A. 1994, "Autoproteolysis in hedgehog protein biogenesis", Science, 266, 1528-1537.
Lenhoff, H. M. 1991, "Ethel Browne, Hans Spemann, and the discovery of the organizer phenomenon", Biological Bulletin, 181, 72-80.
Levin, M. & Mercola, M. 1998, "Gap junctions are involved in the early generation of left-right asymmetry", Developmental Biology, 203, 90-105.
Levin, M., Pagan, S., Roberts, D. J., Cooke, J., Kuehn, M. R., & Tabin, C. J. 1997, "Left/right patterning signals and the independent regulation of different aspects of situs in the chick embryo", Developmental Biology, 189, 57-67.
Lipson, H. & Pollack, J. B. 2000, "Automatic design and manufacture of robotic lifeforms", Nature, 406, 974-978.
Ludwig, W. 1932, Das Rechts-Links-Problem im Tierreich und beim Menschen, Berlin: Verlag Julius Springer.
Mazziotti, M. V., Willis, L. K., Heuckeroth, R. O., LaRegina, M. C., Swanson, P. E., Overbeek, P. A., & Perlmutter, D. H. 1999, "Anomalous development of the hepatobiliary system in the Inv mouse", Hepatology, 30, 372-378.
McManus, I. C. 1979, Determinants of laterality in man, University of Cambridge: Unpublished PhD thesis.
McManus, I. C. 1984, "The genetics of handedness in relation to language disorder," pp. 125-138 in Advances in Neurology, vol 42: Progress in Aphasiology, F. C. Rose, ed., New York: Raven Press.
McManus, I. C. 1985, Handedness, language dominance and aphasia: a genetic model. Psychological Medicine, Monograph Supplement No.8,
Mercola, M. & Levin, M. 2001b, "Left-right asymmetry determination in vertebrates", Annual Review of Cell and Developmental Biology, 17, 779-805.
Mitchison, N. 1965, pp. 51-55 in Aldous Huxley 1894-1963, J. Huxley, ed., London: Chatto & Windus.
Mittwoch, U. & Kirk, D. 1975, "Superior growth of the right gonad in human foetuses", Nature, 257, 791-792.
Mochizuki, T., Yukio, S., Tsuchiya, K., Shirayoshi, Y., Takai, S., Taya, C., Yonekawa, H., Overbeek, P. A., Hamada, H., & Yokoyama, T. 1998, "Cloning of inv, a gene that controls left/right asymmetry and kidney development", Nature, 395, 177-181.
Morgan, M. J. 1977, "Embryology and inheritance of asymmetry," pp. 173-194 in Lateralization in the nervous system, S. Harnad et al., eds., New York: Academic Press.
Morgan, M. J. 1978, "Author's response", Behavioral and Brain Sciences, 2, 325-331.
Morgan, M. J. & Corballis, M. C. 1978, "On the biological basis of human laterality: II. The mechanisms of inheritance", Behavioral and Brain Sciences, 2, 270-278.
MØller, A. P. & Pomiankowski, A. 1993, "Fluctuating asymmetry and sexual selection", Genetica, 89, 267-279.
MÆller, A. P. & Swaddle, J. P. 1997, Asymmetry, developmental instability and evolution, Oxford: Oxford University Press.
National Portrait Gallery 1994, The Sitwells and the arts of the 1920s and 1930s, London: National Portrait Gallery.
Needham, R. 1973, Right and Left: Essays on dual symbolic classification, Chicago: University of Chicago Press.
Nonaka, S., Tanaka, Y., Okada, Y., Takeda, S., Harada, A., Kanai, Y., Kido, M., & Hirokawa, N. 1998, "Randomisation of left-right asymmetry due to loss of nodal cilia generating leftward flow of extraembryonic fluid in mice lacking KIF3B motor protein", Cell, 95, 829-837.
Okada, Y., Nonaka, S., Tanaka, Y., Saijoh, Y., Hamada, H., & Hirokawa, N. 1999, "Abnormal nodal flow precedes situs inversus in iv and inv mice", Molecular Cell, 4, 459-468.
Oppenheimer, J. M. 1974, "Asymmetry revisited", American Zoologist, 14, 867-879.
Oster, J. 1971, "Varicocele in children and adolescents", Scandinavian Journal of Urology and Nephrology, 5, 27-32.
Palmer, A. R. 2000a, "Quasireplication and the contract of error: Lessons from sex ratios, heritabilities and fluctuating asymmetry", Annual Review of Ecology and Systematics, 31 , 441-480.
Palmer, A. R. & Hammond, L. M. 2000b, "The emperor's codpiece: A post-modern perspective on biological asymmetries", International Society for Behavioural Ecology Newletter, 12 (2), 13-20.
Palmer, A. R. & Strobeck, C. 1986, "Fluctuating asymmetry: measurement, analysis, patterns", Annual Review of Ecology and Systematics, 17, 391-421.
Palmer, A. R. & Strobeck, C. 2001a, "Fluctuating asymmetry analyses revisited", Manuscript submitted.
Parraudeau, M., Scott, J., Walsh, C., Oakley, C., Bloom, S., & Brooks, D. 1994, "Late presentation of Kartagener's syndrome", British Medical Journal, 308, 519-521.
Policansky, D. 1982, "The asymmetry of flounders", Scientific American, 246 (May), 96-102.
Ruiz-Trillo, I., Riutort, M., Littlewood, D. T. J., Herniou, E. A., & Baguña, J. 1999, "Acoel flatworms: Earliest extant Bilaterian metazoans, not members of platyhelminthes", Science, 283, 1919-1923.
Ryan, A. K., Blumberg, B., Rodriguez-Esteban, C., Yonei-Tamura, S., Tamura, K., Tsukui, T., de la Pena, J., Sabbagh, W., Greenwald, J., Choe, S., Norris, D. P., Robertson, E. J., Evans, R. M., Rosenfeld, M. G., & Izpisua Belmonte, J. C. 1998, "Pitx2 determines left-right asymmetry of internal organs in vertebrates", Nature, 394, 545-551.
Samatey, F. A., Imada, K., Nagashima, S., Vonderviszt, F., Kumasaka, T., Yamamoto, M., & Namba, K. 2001, "Structure of the bacterial flagellar protofilament and implications for a switch for supercoiling", Nature, 410, 331-337.
Server, P. G., Garcia, H. J., Prieto, G. A., Rigabert, M. M., & Perez, A. M. 1998, "[Right varicocele as first manifestation of situs inversus]", Actas.Urol.Esp., 22, 528-530.
Siewert, A. 1904, "Uber einem fall von Bronchiektasien bei einem Patienten mit situs inversus viscerum", Klinische Wochenschriften, 41, 139-141.
Sitwell, O. 1945, Left hand, Right hand!, London: Macmillan.
Sitwell, O. 1946, The scarlet tree, London: Macmillan.
Spemann, H. 1938, Embryonic development and induction, New Haven: Yale University Press.
Splitt, M., Wright, C., Sen, D., & Goodship, J. 1999, "Left-isomerism sequence and maternal type-1 diabetes", Lancet, 354, 305-306.
Supp, D. M., Brueckner, M., Kuehn, M. R., Witte, D. P., Lowe, L. A., McGrath, J., Corrales, J., & Potter, S. S. 1999, "Targeted deletion of the ATP binding domain of left-right dynein confirms its role in specifying development of left-right asymmetries", Development, 126, 5495-5504.
Supp, D. M., Potter, S. S., & Brueckner, M. 2000, "Molecular motors: the driving force behind mammalian left-right development", Trends in Cell Biology, 10, 41-45.
Tazelaar, M. A. 1929, "The effect of a temperature gradient on the early development of the chick", Quarterly Journal of Microscopical Science, 72, 419-446.
Tazelaar, M. A., Huxley, J. S., & De Beer, G. R. 1930, "Some further effects of an antagonistic temperature gradient upon the frog's egg", Anatomical Record, 47, 1-11.
Ticho, B. S., Goldstein, A. M., & Van Praagh, R. 2000, "Extracardiac anomalies in the heterotaxy syndromes with focus on anomalies of midline-associated structures", American Journal of Cardiology, 85, 729-734.
Tihen, J. A., Charles, D. R., & Sippel, T. D. 1948, "Inherited visceral inversion in mice", J.Hered., 39, 29-31.
Twitty, V. C. 1928, "Experimental studies on the ciliary action of amphibian embryos", Journal of Experimental Biology, 50, 319-344.
Twitty, V. C. 1966, Of scientists and salmanders, San Francisco: W H Freeman.
Vinken, P. 1999a, The shape of the heart, Amsterdam: Elsevier.
Von Kraft, A. 1999, "Symmetry and asymmetry in the development of inner organs in parabiotic twins of amphibians (Urodela)", Laterality, 4, 209-255.
Wagner, M. K. & Yost, H. J. 2000, "The roles of nodal cilia", Current Biology, 10, R149-R151.
Wagner, R. 1888, Gesammelte Schriften und Dichtungen von Richard Wagner, volume 6, 2 edn, Leipzig: G.M. Fritzsch.
Wehrmaker, A. 1969, "Right-left asymmetry and situs inversus in Triturus alpestris", Wilhelm Roux' Archiv für Entwicklungsmechanik, 163, 1-32.
Weismann, A. 1892, Das Keimplasma. Eine Theorie der Vererbung., Jena: Gustav Fischer.
Whitlock, M. 1996, "The heritability of fluctuating asymmetry and the genetic control of developmental stability", Proc.R.Soc.Lond.B Biol.Sci., 263, 849-854.
Whitman, M. & Mercola, M. 2001c, "TGF-b superfamily signalling and left-right asymmetry", Science's STKE, http://www.stke.org/cgi/content/full/OCV_sigtrans;2001/64/re1.
Wolpert, L., Beddington, R., Brockes, J., Jessell, T., Lawrence, P., & Meyerowitz, E. 1998, Principles of Development, London: Current Biology.
Worboys, M. 1981, "The British Association and Empire: Science and social imperialism, 1880-1940," pp. 170-187 in The Parliament of science: The British Association for the Advancement of Science, 1831-1981, R. MacLeod & P. Collins, eds., Northwood, England: Science Reviews.
Wright, C. V. E. 2001, "Mechanisms of left-right asymmetry: What's right and what's left?", Developmental Cell, 1, 179-186.
Yokoyama, T., Copeland, N. G., Jenkins, N. A., Montgomery, C. A., Elder, F. F. B., & Overbeek, P. A. 1993, "Reversal of left-right asymmetry - a novel situs inversus mutation", Science, 260, 679-682.
Yost, H. J. 2001, "?????????", International Review of Cytology, 203, 357-381.
Ziegler, P. 1999, Osbert Sitwell: A biography, London: Pimlico.